
Cladistics is a method of reconstructing phylogeny without necessary recourse to either fossils or molecular distances. It is required to trace the evolutionary changes within a group of organisms (a series of taxa), and the following stages are followed.
First, one or more outgroups are selected; these are taxa that are known to be outside the group under study (the ingroup), but not too distantly related to it. Second, the characters that vary among taxa of the ingroup are listed. Third, the character states (the ways in which the ingroup taxa differ) are listed as objectively as possible, and the corresponding states of the outgroup(s) are also listed. The states shared by the outgroup with members of the ingroup are deemed to be most plausibly inherited from their common ancestor: They are called symplesiomorph, or shared primitive. States that occur only in one or more members of the ingroup and not with the outgroup(s) are deemed to have evolved since the ingroup came into being: They are called apomorph, or derived. Some of these apomorph states are confined to a single member of the ingroup: These are called autapomorph, or uniquely derived. Other apomorph states are shared between different members of the ingroup: These are called synapomorph, or shared derived. It is the synapomorph states, of course, that are evidence for phylogenetic relatedness.
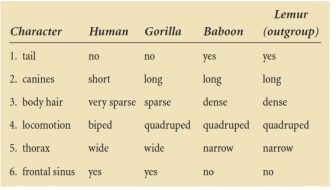
The simplest case reconstructs the phylogeny of just three ingroup members (the three-taxon problem). An example in anthropology might be the reconstruction of the phylogenetic relationships of human, gorilla, and baboon. We might take the lemur as a sensible outgroup; it is acknowledged that lemurs are phylogenetically more distant from the members of the ingroup than they are from each other. We can list a few characters that vary in the ingroup, and their states in the different taxa, as follows:
In Characters 2 and 4, human is different from the outgroup, but uniquely so: The states are autapo-morph (uniquely derived). These two characters therefore show how unusual human is, but not which of the other two ingroup members it more resembles. But if we take Characters 1, 5, and 6, we see that though human is again different from the outgroup, it shares the derived states with gorilla; these states are therefore synapomorph (shared derived) and are evidence that human and gorilla are phylogenetically closer to each other than to baboon. Finally, Character 3 is a special case; it has three states, not just two, and logically, the gorilla state (sparse) is intermediate between that of baboon and the outgroup (dense) and that of human (very sparse).
 The deduced primitiveness or derivedness of the character states is called their polarities. The character states are typically coded; in the above case, all the characters have States 1 and 2, except for Character 3, which has States 1, 2, and 3. One then has the option of whether State 2 is to be treated as intermediate between 1 and 3 (ordered) or whether all three states are equidistant.
The deduced primitiveness or derivedness of the character states is called their polarities. The character states are typically coded; in the above case, all the characters have States 1 and 2, except for Character 3, which has States 1, 2, and 3. One then has the option of whether State 2 is to be treated as intermediate between 1 and 3 (ordered) or whether all three states are equidistant.
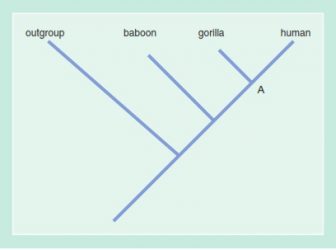
We arrive, then, at the phylogeny shown in this figure:
At Node A, the human/gorilla synapomorph states of Characters 1, 3, 5, and 6 appear. Between Node A and human, the human autapomorph states of Characters 2, 3, and 4 appear.
We refer to gorilla and human as a clade (branch) with respect to baboon and the outgroup, while of course gorilla, human, and baboon together form a clade. Human and gorilla are sister groups (or if they are species, sister species); the human-plus-gorilla clade is the sister group of baboon.
We can then add other taxa to the analysis: chimpanzee, orangutan, gibbon, for example. The more taxa we add, the more unwieldy the analysis becomes.
What renders it yet more difficult is the existence of homoplasy. This is where an evolutionary event occurs more than once. The three types of homoplasy are:
- Convergence: two rather different taxa independently evolve the same character state
- Parallelism: the same character state evolves independently in two descendants of the same ancestor
- Reversal: a more apomorph state, after evolving, is lost again
Homoplasy can easily be mistaken for homology (the unitary occurrence of an event); when two taxa share an apomorphic state, generally the only way to deduce whether they share it by homology or by homoplasy is by parsimony. On the assumption that an evolutionary change is a rare event, an apomorphic state is more likely to have occurred once than twice or more. The cladist will count up the apparent synapomorphic states between taxa A and B, and those between A and C and between B and C, and will usually suppose that the two that share the largest number are the sister groups. The smaller numbers of shared states that are incompatible with this conclusion are deemed to constitute homoplasy.
When a large number of taxa are being analyzed, using a large number of characters, then cladistic analysis simply cannot be performed without the aid of a computer. There are several computer packages available to perform cladistics quickly and efficiently, giving the number of steps (evolutionary changes) and several indices that record the amount of homoplasy (consistency index, retention index, and so on). When the true phylogeny is already known, or suspected, most of these computer packages can compare the analyzed phylogeny against the known one and so test the assumption of parsimony. In some cases, the result has revealed that homoplasy is unexpectedly frequent.
There is no reason why fossil taxa should not be included in a cladistic analysis. The results often have a bearing on the potential ancestral position of a given fossil, although it is important to appreciate that the function of a cladistic analysis is to discover sister group relationships, irrespective of whether one of a pair of sister groups might actually be an ancestor.
The assumption behind character counting is, of course, that each change of state represents a mutation. If there is reason to think that a state change is more complex than this, then different characters can be given different weights, so that one counts more in an analysis, and another less. Finally, molecular data can be analyzed cladistically as well as by distance methods. The subject of cladistics is a complex one, with its own language and methodologies. Its influence on evolutionary anthropology has been enormous. A separate question is whether the results can be directly plugged in to make a taxonomic scheme or whether taxonomy should be more than simply a representation of phylogeny.
Clans
The anthropological term clan comes from Gaelic clann and Old Irish clann or cland, denoting a group claiming descent from a common ancestor. The word was extended to refer to similar kinds of groups in other cultures at least from the early 16th century, and by the 1860s it had entered into sociological and anthropological usage. Anthropologists have given a number of different labels to groups recruited by filiation or descent: for example, gens (plural gentes) by Morgan and sib by Lowie, who reserved gens for the patrilineal form and clan for the matrilineal form. Clan has become the preferred term, however.
In anthropological usage, a clan is a social group whose members share a doctrine of common ancestry but who do not necessarily trace descent from a common ancestor by known genealogical links. A clan may consist of more than one lineage whose members trace descent from a common ancestor by known (but possibly fictive) genealogical links. The group may be recruited by patrifiliation to form a patrilineal clan, by matrifiliation to form a matrilineal clan, or by a combination of these to form a cog-natic clan. Adoptive links may be important. There may be a difference between the descent ideology and actual practice, as in parts of Papua New Guinea, where a matrifilial link becomes converted to a patrifilial one.
Clans may be exogamous; that is, a member ought to marry someone from a clan other than his or her own. Clan organization may articulate with other forms: Two or more clans may cluster into phratries; clans may be distributed between two moieties; or clans may be distributed between four semimoieties. A number of clans may form a larger entity, such as a phratry, tribe, or language group. This hierarchical structure of identities is most elaborate in models of segmentary lineage systems applied to societies of the southern Sudan.
Societies vary in the predominant lineality of clans; in some societies, patrilineal or patrifilial groups are predominant, while in others (though more rarely) matrilineal groups are the most common form. Some societies combine patrilineal and matrilineal clans in a system of double descent. Groups in Polynesian societies draw on ambilineal or cognatic descent to form groups in which an individual has potential claims in a number of groups, through both parents and four grandparents, although only one claim is likely to be activated.
Clans and other kinds of groups coexist with kinship, which is normally bilateral. The form of kin classification, however, may be strongly shaped to fit lineage and moiety identities. In the case of unilineal groups, each individual is often connected to the other parent’s clan through complementary filiation, yielding rights that are not transmitted to the next generation.
In general, clan organization has been found in societies in which kinship and descent have provided the main bases for the organization of society, although it can be combined with hierarchical forms of organization. It has persisted in many areas with colonial and postcolonial state structures.
Clans and Hierarchy
Clan organization is capable of elaboration to incorporate various forms of hierarchy. According to the model of the conical clan, which has been applied to descent groups in parts of Polynesia, the birth order of ancestors ideally determines the rank order of their descendant lineages, although a good deal of manipulation is possible, especially where these groups are ambilineal or cognatic. Raymond Firth applied the term ramage to the ambilineal Polynesian clan. Hunter-gatherer societies of the northwest coast of North America were hierarchical. Among coastal Tshimshian, for example, the four matrilineal clans were ranked in order, and members of each clan were ranked, with chiefs, who possessed honorific titles, at the top of the rank order.
In some chiefdoms, royal clans and lineages were distinguished from commoner ones. Among Bantu peoples of southern and central Africa, such as the Zulu, Swazi, and Bemba, the descendants of a chief’s clan form a ruling caste, and clans are ranked according to the tradition of original migration into the area or the degree of relationship with the chief’s descent group.
Clans in Social Theory
The concept of clan has featured in general theories of social organization. For example, the 19th-century theory of totemism outlined an ideal type of “primitive” society in which each exogamous clan was linked to a totemic species. In his outline of the evolution of the division of labor, Emile Durkheim contrasted “segmental structure” associated with a minimal division of labor and “mechanical solidarity,” with a more generalized structure of permeable groups associated with a greater division of labor and “organic solidarity.”
The concept of clan became linked with the concepts of corporation and corporate group, deriving from the legal history of Henry Maine and Max Weber’s theory of the verband, but with its ultimate origins in Western law of incorporation. A corporate clan is thus a descent group whose identity persists beyond the life of any individual member and that holds some kind of property as a group, such as land and/or totemic insignia. In the structural functionalist anthropology of Africa, the clan became one variety of the unilineal descent group, theorized by anthropologists such as Meyer Fortes. Thus, in structural functionalist anthropology, “clan” and related concepts were linked to descent theory, in which descent was seen as a major component of social structure. This approach contrasted with alliance theory, associated with Claude Lévi-Strauss, in which groups such as clans were linked in various ways by marriage exchange.
Critiques of the Concept
From the early 1960s, cultural and social anthropologists, including John Barnes, Robert Barnes, Ian Keen, Adam Kuper, and Roy Wagner, have criticized lineage theory and the use of concepts such as clan and sib on the grounds that the assumptions and metaphors implicit in the concepts distort ethnographic reality. The concept of clan is perhaps best thought of as a model that approximates the constitution of social identities and their entailments to varying degrees.
References:
- Kuper, A. (1982). Lineage theory: A critical retrospect. Annual Review of Anthropology, 11, 71-95.
- Wagner, R. (1974). Are there social groups in the New Guinea Highlands? In M. Leaf (Ed.), Frontiers of anthropology (pp. 95-122). New York: Van Nostrand.
