
Among living primates, only humans are bipedal. It is not certain when this unique feature emerged, but it must have been before the 3.6 million-year-old Laetoli footprints were made. Although the prints were not made by completely modern feet, they are unequivocally the prints of bipeds. They are the impressions of feet that lacked a distinctive human rounded ball, or swelling, at the base of the great toe, that had no well-defined arch, and that retained ever so slightly divergent great toes. Somewhat later in time, the well-known 2.9 million-year-old Australopithecus afarensis Lucy fossil is the earliest human ancestor to display the clear skeletal hallmarks of bipedalism. Earlier fossils are either not yet described or lack the two most diagnostic parts, the pelvis and the distal (i.e., lower) femur.
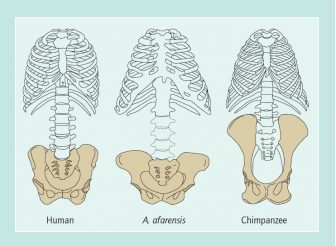
Despite the paucity of fossils near the beginning of the human lineage, most paleontologists regard bipedalism as the earliest distinctively human feature to have evolved in hominins. Hominin refers to members of the taxonomic family Homininae, which consists of human ancestors and collateral species after the lineage split from that leading to chimpanzees. In great apes, the pelvis is vertically elongated so that it makes up much of the back (see figure), leaving the back somewhat inflexible. The human pelvis is vertically short and shaped like a bowl with an open bottom, the opening being the pelvic inlet or birth canal. Humans also possess a valgus femur, a femur that angles in at the knee, whereas ape femora have no angle, giving them a bowlegged appearance. A. afarensis clearly exhibits these two hallmarks of bipedalism. Many other early hominin features, however, are more primitive. Early hominins have a partly divergent great toe, long curved lateral toes, short legs, a knee joint that is small and flexible, a cone-shaped rib cage, long robust arms, disproportionately long forearms, a scapula or shoulder blade with a tilted-up joint surface, and curved robust fingers with distinctive concavities on the inner surface to accommodate large finger flexor tendons.
Two features in A. afarensis are neither chimpanzee-like nor humanlike. First, the pelvis, while human shaped, is extraordinarily broad. Second, the joint at the base of the great toe, deep in the foot near the ankle, is shaped like a hinge. As with a hinge, it allows little movement up or down and is therefore a rigid strut for pushing off during walking or running, just as is the great toe of humans. Like a hinge, it swings side to side, perhaps to allow the great toe to grip small branches. In chimpanzees, the whole foot is modified so that the great toe functions like a human thumb.
 Disagreement among experts on the locomotor habits of early hominins has little to do with these details themselves and much to do with their interpretation. Some argue that the apelike features are holdovers from an arboreal ancestor, “primitive retentions,” and therefore unimportant for inferring behavior. Others interpret the primitive features more straightforwardly.
Disagreement among experts on the locomotor habits of early hominins has little to do with these details themselves and much to do with their interpretation. Some argue that the apelike features are holdovers from an arboreal ancestor, “primitive retentions,” and therefore unimportant for inferring behavior. Others interpret the primitive features more straightforwardly.
As might be expected, given that experts disagree on function, the circumstances that led to the evolution of bipedalism are quite contentious. The first bipedalism origin hypothesis was that of Charles Darwin. Although some human fossils had been discovered in his lifetime, Darwin never mentioned them. Rather, he compared living humans to apes and suggested that our enormous brains and upright locomotion were linked. Quite the opposite of our current understanding, Darwin suggested that humans evolved large brains when still apelike tree dwellers. Having conceived of tools, they became bipedal to free the hands to carry and manipulate tools, and having adopted tool use, lost their projecting canines. The fossil record shows us this is backward. Human-sized brains evolved only in the last tenth of our existence, whereas bipedalism evolved early.
In the early 1900s (1912, 1923), Sir Arthur Keith observed that gibbons held their legs extended during their distinctive hand-over-hand suspensory locomotion, called brachiation. He suggested that humans were the product of a trend for progressively more erect posture in primates, starting with a rather typical quadrupedalism in monkeys to a more erect quadrupedalism in apes, an erectness required by their long forelimbs and culminating in bipedal humans. He offered no argument for why such an unwavering progression was inevitable, and current theory does not consider such evolutionary inertia possible. Keith noted that gibbons had large muscles of the upper limb but small lower back muscles. Large back muscles are used by monkeys to uncoil the back when leaping. He found more than 80 shared by apes and humans that are not shared with other primates. As Keith’s hypothesis matured, gibbons, great apes, and human ancestors were all seen as sharing long arms, rotatable wrists, mobile shoulders, short backs, broad torsos, pronounced shoulders, and powerful upper bodies, all as adaptations to brachiation.
Although Keith’s scenario was not uncontested, many saw Darwin’s tool-carriage hypothesis as easily melded with Keith’s increasing-erectness scenario to model bipedalism having evolved as a brachiator included more tool use in its daily behavior. Just as the Keith perspective achieved this splendid internal consistency, contrary evidence emerged to reinvigo-rate the opposition. At mid-century, Louis S. B. Leakey brought his considerable persuasiveness and the first Proconsul fossils to bear on the issue, weighing in against the Keith brachiationist model. Proconsul, an early ape—too early to inform us on the anatomy of the chimp-human common ancestor, we now know— had a monkeylike body. Bolstering Leakey’s arguments was the steady discovery of hominin fossils at Sterkfontein, South Africa, hominins with unsettlingly ape-sized brains.
Although carrying/tool use hypotheses bore up rather well in the face of this contrary evidence, by the 1960s a multitude of bipedalism-origin contenders crowded the field. The tool-use hypothesis was embellished to suggest that throwing in particular was critical, though this did nothing to resolve the brain-size/tool-use problem. Others suggested that bipedalism might have freed the hands for carrying something other than tools—infants, for instance. This certainly touches an important evolutionary pressure point; however, no other primate infant needs help clinging to its mother. Carrying food to a home base to provision mates or other group members was proposed in the early 1960s and found its adherents.
Raymond Dart, famous for his hunting hypothesis for human origins, saw bipedalism as having evolved to allow long-distance viewing over tall grasses, to spot prey in Dart’s view, to spot predators in the view of others. Such vigilance bipedalism is not extremely rare among chimpanzees, nor have other mammals adopted bipedalism due to predation pressure. Noting that chimpanzees are bipedal during social displays, and that bipedalism makes individuals appear larger, some suggested that bipedalism functions to frighten predators or to bully competitors. Threats, however, are more effective when rare, the better to contrast with usual behavior. Surely low-ranking bipedal bluffers would attract aggression from dominants.
The 1960s also saw the first presentation of the aquatic hypothesis, the idea that bipedalism evolved as a wading posture for collecting aquatic foods. Critics argued that natural buoyancy reduces the need to evolve anatomical adaptations to this sort of bipedalism, and they pointed out that other aquatic mammals are not bipedal.
In 1970, Cliff Jolly proposed that early hominins stood erect to gather grass seeds, a seed-eating hypothesis. He argued that small seeds required a constant two-handed picking action to achieve a reasonable rate of ingestion; bipedalism freed the hands. The hypothesis was an immediate hit, but criticisms gradually mounted. Grasses can be pulled over and harvested without bipedalism. The hominin habitat was more likely woodland or forest, rather than grassland. Hominin dental microwear suggested a diet of fruits rather than seeds. As the hypothesis was debated, newly discovered A. afarensis fossils exhibited more arboreal anatomy than expected. Mike Rose suggested a saving modification. He had observed that baboons were bipedal when harvesting small fruits from bushes. Perhaps small fruits rather than seeds were picked bipedally. This friendly amendment to the Jolly perspective yielded the small-object feeding hypothesis, salvaging Jolly’s most appealing arguments while addressing their weaknesses. Richard Wrangham argued that when trees are packed together closely, bipedal shuffling uses less energy than repeatedly lowering and raising the torso to walk quadrupedally. Most important, this suggested a means by which bipedal locomotion might gradually increase in frequency, even if inefficient.
In 1980, Peter Rodman and Henry McHenry drew on data showing that human bipedalism was efficient compared to chimpanzee quadrupedalism to suggest that bipedalism arose to make moving between increasingly dispersed feeding trees more efficient. Critics pointed out that some quadrupeds are more efficient walkers than humans, suggesting that bipedalism was not a likely adaptation when efficiency is desired. A quadrupedal hominin ancestor, moreover, would lose efficiency by standing up and adopting a locomotion for which it was not adapted. Although the authors had claimed that bipedalism was efficient only in relation to apes, many saw an initial inefficiency, should a quadruped stand up, as an insurmountable evolutionary barrier.
In 1980, Owen Lovejoy gave the carrying hypothesis a complete overhaul. He emphasized that bipedal locomotion was a ridiculous adaptation. Its two points of contact rather than four afford less maneuverability, less stability, and less speed. Unlike other mammals, bipeds cannot use their backs to increase their stride, and with only two limbs and therefore half the muscle power to move the body, humans accelerate slowly and stay slow. Lovejoy set high hurdles barring the evolution of bipedalism and then found a way over them: He suggested that bipedalism dramatically increased female reproductive rates. He proposed that males who collected and carried supplementary food to females could significantly increase their birthrates. If males had high confidence in paternity, they would aid their own offspring. As Lovejoy envisioned it, bipedalism, monogamy, lack of visible estrus (sexual swellings), terrestriality, and the relative success of humans as a species were interrelated.
This elegant hypothesis was at odds, critics argued, with great body size differences between males and females. In most monogamous species, males and females are similar-sized. Long curved toes, short legs, and especially wide hips imply a rather inefficient bipedalism, whereas carrying would seem to require efficiency. Others pointed out that tooth morphology suggested that australopiths (“australopithecines” in traditional taxonomy) were fruit eaters, and carrying fruits when the arms are needed to gather more food is clumsy, if not impossible. The obvious solution to this problem, a carrying device, obviates the need for bipedalism. Furthermore, provisioning need not mean carrying; ingestion and regurgitation accomplishes the task without the disadvantages of bipedal-ism. Others noted that long male absences while gathering would prevent males from guarding females from other males, a behavior common in monogamous species.
Lovejoy responded that large body size in males might be related to the larger day-ranges required for provisioning, that body size differences have been overstated, that arboreal traits are merely holdovers from a previous arboreal lifeway, and that many of the arboreal adaptations were overstated anyway.
Peter Wheeler suggested that bipedalism evolved to avoid overheating. He noted that a bipedal hominin exposes only 7% of its surface to sunlight, whereas quadrupeds expose 20% of their surface to the sun. Wind speed is greater and temperatures lower only a meter above the level baboons are forced to walk in, he found, and bipeds lift their torsos into this cool zone. Wheeler’s hypothesis was criticized because the reduction in thermal radiation was only significant near noon, when few animals are active anyway. Others noted that early hominin habitats were likely partly wooded, or even forested. Living species adapted to high heat environments are slender to better dissipate heat, whereas the short legs, wide hips, and robust upper bodies of australopiths are stoutly heat retaining. Furthermore, the inefficient locomotion of australopiths would generate considerable heat. Wheeler’s heat-stress hypothesis seems an excellent explanation for the long-legged, slender hipped morphology of the earliest Homo specimen, WT 15,000, but critics argue that it cannot explain the evolution of bipedalism.
In 1994, Kevin Hunt presented data on chimpanzee locomotion and posture that supported a small-object feeding hypothesis, albeit with a more arboreal slant than the classic Jolly and Rose hypothesis. Hunt found that in chimpanzees, most behaviors offered as reasons to adopt bipedalism were rare: display, tool use, carrying, and looking over obstacles made up only 1% or 2% of bipedal behavior each. Feeding made up 85% of bipedal episodes. Chimpanzees were bipedal most often when feeding on small fruits from small trees. Small trees, he noted, were only found in the driest parts of the chimpanzee range. Chimpanzees were bipedal when reaching up to feed from the ground, as Rose and Jolly might have expected, but also when harvesting fruits arboreally. Small trees elicited bipedalism because their small branches were too flexible to sit on, whereas a modified bipedalism where the lateral toes gripped lower branches and a hand gripped an overhead branch was quite stable. He speculated that if prehominins were confined to habitats drier and more open than those in which chimpanzees live, bipedalism might evolve as a feeding adaptation. This interpretation is consistent with Yves Coppens’s argument that hominins were restricted to East and southern Africa by geographic barriers such as the Great Rift, resulting in a dry-habitat ape (hominins) and a moist-habitat ape (chimpanzees).
 Hunt’s analysis of chimpanzee locomotion and posture suggested that curved robust fingers and toes, mobile shoulders, and cone-shaped torsos were evolved for arm hanging. Gripping great toes, short hind limbs, and short stiff backs, he argued, were adaptations to hand-over-hand vertical climbing. Australopiths share arm-hanging traits with chimpanzees, but not climbing adaptations. Because small trees can be ascended with a single agile leap, harvesting fruits from small trees requires none of the vertical climbing anatomy forest-adapted chimpanzees need to access tall trees. He explained that wide hips and short legs in early hominins functioned to increase stability on unstable branches by lowering the body’s center of gravity. Short but curved and robust fingers evolved to grip small branches—the only kind available in these small trees—during arm hanging. A tilted-up shoulder joint in Lucy and a cone-shaped torso reduce stress on the shoulder and torso during one-handed hanging. Curved robust toes functioned to grip branches during arm-hanging-bipedalism (see Figure 2). These arm-hanging features are found in the first hominin specimens over 6 million years old and persist through Australopithecus afarensis, through A. africanus, and right up to the last fossils that cannot be called Homo. None of these arboreal traits are found in Homo erectus, suggesting that arboreal traits disappear quickly once the trees are abandoned.
Hunt’s analysis of chimpanzee locomotion and posture suggested that curved robust fingers and toes, mobile shoulders, and cone-shaped torsos were evolved for arm hanging. Gripping great toes, short hind limbs, and short stiff backs, he argued, were adaptations to hand-over-hand vertical climbing. Australopiths share arm-hanging traits with chimpanzees, but not climbing adaptations. Because small trees can be ascended with a single agile leap, harvesting fruits from small trees requires none of the vertical climbing anatomy forest-adapted chimpanzees need to access tall trees. He explained that wide hips and short legs in early hominins functioned to increase stability on unstable branches by lowering the body’s center of gravity. Short but curved and robust fingers evolved to grip small branches—the only kind available in these small trees—during arm hanging. A tilted-up shoulder joint in Lucy and a cone-shaped torso reduce stress on the shoulder and torso during one-handed hanging. Curved robust toes functioned to grip branches during arm-hanging-bipedalism (see Figure 2). These arm-hanging features are found in the first hominin specimens over 6 million years old and persist through Australopithecus afarensis, through A. africanus, and right up to the last fossils that cannot be called Homo. None of these arboreal traits are found in Homo erectus, suggesting that arboreal traits disappear quickly once the trees are abandoned.
Critics noted that neither chimpanzees nor other primates in dry habitats have evolved bipedalism, that it is not clear that hominins evolved in open habitats, that posture is not stressful enough to demand the wholesale anatomical changes required for bipedalism, and that small fruit-small tree resources may not have been abundant enough for hominins to specialize in them. Hunt responded that evidence that fauna associated with hominins suggests that they were found in drier and more open habitats than chimpanzees, that such habitats have abundant small trees, that bipedal feeding postures require adaptations to prevent fatigue, and that bipedal posture encourages bipedal locomotion.
Although there are unanswered criticisms to each, Wheeler’s thermal radiation avoidance hypothesis, Lovejoy’s provisioning hypothesis, and the small-object postural feeding hypothesis are the three bipedal origins scenarios most often cited by scientists to explain the evolution of bipedalism.
References:
- Coppens, Y., & Senut, B. (Eds.). (1993). Origine(s) de la Bipedie chez les Hominides. Paris: CNRS Cahiers de Paleoanthropologie.
- Stanford, C. (2003). Upright: The evolutionary key to becoming human. Boston: Houghton Mifflin.
- Strasser, E., Fleagle, J. G., McHenry, H. M., & Rosenberger, A. (Eds.). (1998). Primate locomotion: Recent advances. New York: Plenum.
