
The study of locomotor adaptations is fundamental to the study of primate evolution and ecology. The primates are particularly interesting because they manifest a remarkable diversity of locomotor styles. While workers have often focused on masticatory morphology, data on patterns of locomotion also provide information on the niches occupied by recent and fossil species. While some other mammalian orders contain arboreal, semiterrestrial, and terrestrial representatives, few show the diversity of locomotor modes that are exhibited in the primates. Arboreal primates include various types of quadrupeds, brachiators, “quadrumanus” climbers, and vertical clingers and leapers. Terrestrial primates include quadrupedal forms and the unique striding bipedal hominins.
While considerable attention has been devoted to the unique and spectacular forms of primate locomotion, such as brachiation and vertical clinging and leaping, the most common locomotor mode received comparatively little attention until recently. Most primate species are quadrupedal, and the quadrupedal species often bridge the gap between locomotion on the ground and in the arboreal milieu. The assumption that all primate quadrupeds are basically the same in their anatomy and behavior is inaccurate because of both phylogenetic and ecological considerations. In a phylogenetic sense, quadrupeds are represented in a variety of infraorders, including Lemuriformes, Chiromyiformes, Loriformes, and Simiiformes. Within the Simiiformes, the Platyrrhini include clawed forms and other species with prehensile tails. In contrast, there are no known catarrhines with either claws or prehensile tails. Within the various primate families, a diversity of ecological niches are occupied.
John and Prue Napier characterize quadrupedal primates as using fore- and hindlimbs together during walking and running. They have differentiated generalized quadrupeds and “semibrachiators”: In the former, arms and legs contribute about equally to locomotion; in the latter, the forelimbs are supportive, and the hind limbs are propulsive. Michael D. Rose points out that the term quadrupedalism can be used in two ways. It can be used to describe locomotor activity in which the animal stands and moves on all four limbs, locomotion is on horizontal or almost horizontal supports, and locomotion occurs above the support surface (in distinction to suspension). Rose also states that the term is used in a broader sense—to refer to a locomotor repertoire that has a major element of quadrupedal behavior in it.
Cercopithecoids as Models to Study Quadrupedalism
Because of focus on the evolution of extant hominoids from a Miocene stock, much of the detailed anatomical work and naturalistic study has focused on those species that have been characterized as brachiators or semibrachiators: apes and certain New World monkeys and Old World colobines.
The most taxonomically diverse and numerically successful nonhuman catarrhines are Old World monkeys. It is unfortunate that Adoph Schultz’s generalization, that all cerecopithecoids are similar in their postcranial morphology, has had considerable influence. Cercopithecoids have been considered to be a monolithic block of pronograde quadrupeds, not showing the adaptations that distinguish certain species of New World monkeys and basically similar in their locomotor behavior and postcranial structure. Because of the fundamental assumption of similarity in the postcrania of cercopithecoids, with the exception of studies that contrast colobines with cercopithecines, there has been a tendency to emphasize or assume similarities of fossil cercopithecoids to their closest living relatives. An example of this is the reconstruction of Theropithecus oswaldi as a digitigrade baboon without specific study of the hand or wrist bones. In addition, data from derived from experimental research on quadrupedal representatives of other orders (such as opossums and cats) have been uncritically applied to quadrupedal cercopithecoids.
These approaches mask the variation that exists within and between cercopithecoid species. One comparison that has been made is between colobines and cercopithecines, in part because of the leaf-eating habits of colobines, the reduction of the thumb in African species, and the discovery of fossil forms such as Paracolobus; with the exception of “semi-terrestrial” forms such as langurs and the unusual fossil Cercopithecoides, the assumption has been that colobines should show features that are adapted to a more arboreal set of niches than the cercopithecines. While the comparison is useful, the taxonomic emphasis conceals fundamental differences between monkeys within the colobines and cercopithecines: that is, between palmigrade versus digitigrade types and between the more arboreal versus more terrestrial forms.
Since 1980, I have been engaged in a project that focuses on the locomotion of African cercopithecoids, especially extant Cercopithecus aethiops and extinct Theropithecus oswaldi, to answer questions of proximate and ultimate causation of quadrupedal behavior and anatomy. Cercopithecoids are ideal subjects because they are more “generalized” in their quadrupedal behavior than are species with specializations such as prehensile tails.
Contrary to the recent statement by Alan Turner and Mauricio Anton, in Evolving Eden: An Illustrated Guide to the Evolution of the African Large-Mammal Fauna, that vervet monkeys are always palmigrade in their locomotion, my own observation of vervets in the field and captivity demonstrates that some individuals are digitigrade while others are palmigrade; the precise distribution of this behavioral difference in the field has not yet been determined. The dichotomy would be considered unusual intraspecifically, because most species appear to manifest one hand posture or the other, but not both. It is an especially important control because behavioralists and morphologists are concerned with the effects of phylogenetic constraints and fabricational factors. In any study employing interspecific or intergeneric comparisons, there is an element of doubt whether the observed differences reflect differences in ways of life (niche) or whether they are due to phylogenetic or fabricational factors. One way of increasing the likelihood of correlating differences in form with differences in function is to perform intraspecific comparisons of populations in different habitats. Examples of this approach include K. A. Monk’s study of grasshoppers, B. D. Patterson’s work on chipmunks, S. L. Linsay’s research on Douglas squirrels, and R. F. Johnston’s classic investigation of European sparrows. In the research on vervets, the problem of phylogeny is controlled to a great degree because much of the work is done with representatives of different local groups of the same species.

Quadrupedal locomotion in vervet monkeys on Lake Naivasha, Kenya. Field observation demonstrated that individuals were consistently palmigrade in hand posture. They actively foraged on the ground (left) and in the trees. In the terrestrial context, foodplants such as Amaranthus hybridus, Oxygonum sinuatum, Sida cuneifolia, Cynodon dactylon, and Opuntia were routinely consumed. In the arboreal context, the fruit of Ficus praetoriae and F. wakefieldii were eaten. Acacia xanthophloea provided gum, leaf buds, young leaves, flowers, seeds, and thorn bases; most of these items were consumed in the terminal branch setting but gum and red mites were eaten from the trunk and boughs (middle). The terminal branch setting provided extensive lateral contact between crowns, producing virtually continuous arboreal pathways for the monkeys (right) in their core area. There was seasonal variation in the consumption of some foods (e.g., Acacia flowers), which affected the amount of time spent terrestrially versus arboreally, while other foods (e.g., Opuntia pears and Acacia gum) are consumed during all seasons. While the Naivasha vervets ate at least 40 different species of plants, they were able to consume various items from yellow fever trees throughout the year.
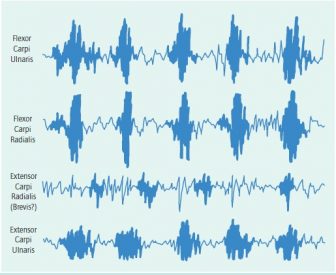
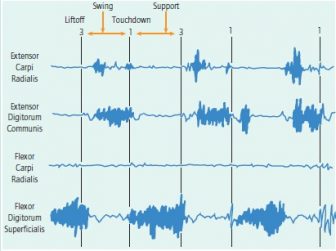
Comparison of two long flexors and two extensors of the wrist in the male palmigrade-digitigrade hybrid. Note in-phase activity of the flexors and extensor carpi ulnaris during the support phase of quadrupedal walking. There is a discrete burst of activity of extensor carpi radialis (brevis?) that is generally out-of-phase with the other muscles.
The aim of the research has been to correlate variations in hand-posture behavior (digitigrady versus palmigrady) and in underlying morphology, with variations in local troop habitat structure and substrate preference. The proposition that is being tested is that digitigrady is correlated with habitual terrestriality and open habitat, while palmigrady is correlated with habitual arboreality and more closed habitat. I am therefore seeking to determine
(1) whether there is a predictable relationship between behavior, morphology, and ecology;
(2) whether certain kinds of African environments impose constraints on the adaptations of monkeys; and (3) whether these are represented simultaneously in their ecology and morphology. Study of forelimb anatomy of fossil Theropithecus oswaldi provides a perspective as to whether some characters may be size related in their expression.
My research has a number of components (1) field study of the locomotor behavior and habitat of a palmigrade group on Lake Naivasha, Kenya, for comparison to a later study of digitigrade vervets; (2) trapping of individuals from the group that was studied in the field for electromyographic and cineradiographic comparison of palmigrady with digitigrady; and (3) osteological comparison of the forelimbs of species of African cercopithecoids, which have been described in the literature as either habitually palmigrade or digitigrade, examination of the range of variability within C. aethiops, and relation of intraspecific to interspecific differences between palmigrade and digitigrade forms. The results of all of these approaches would be too extensive to enumerate here, but some results of the electromyo-graphic and cineradiographic in vivo experiments on primate quadrupedalism are presented below.
Experimental Study of Digitigrady and Palmigrady During Quadrupedal Locomotion
Electromyography was accomplished on two vervet monkeys and an anubis baboon. One vervet was a palmigrade adult female from Lake Naivasha. The second was a subadult male, born at the Institute for Primate Research, National Museums of Kenya. The male was a product of a cross between a digitigrade male vervet and a palmigrade female. The baboon was a late infant-early juvenile, trapped in the wild but obtained from a commercial primate importer, and was consistently digitigrade in hand posture. The research was accomplished in the facility of the Department of Anatomical Sciences, State University of New York at Stony Brook, with the kind permission and cooperation of Jack T. Stern, William Jungers, and Susan Larson. The muscles that were investigated were flexor carpi ulnaris, flexor carpi radialis, extensor carpi radialis (brevis in the vervet and longus in the baboon), extensor carpi ulnaris, extensor digitorum communis, flexor digitorum superficialis (mainly to the third and fourth rays), and flexor digitorum profundus (mainly to the third and fourth rays). Eight experiments produced useful results. Two main questions were addressed: (1) Are the patterns of muscular activity different during support in habitually palmigrade monkeys versus habitually digitigrade monkeys? and (2) are the patterns of muscle activity similar to those documented in knuckle-walking apes?
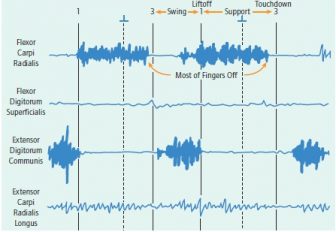
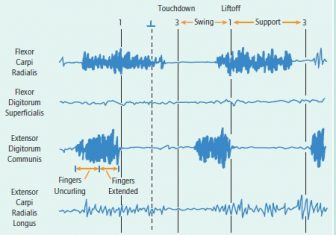
Two sequences of quadrupedal walking in a young male Papio anubis: digitigrade walking (with thumb intermittently contacting surface) on a horizontal branch (top) and fast digitigrade walking on the ground (bottom). Flexor carpi radialis is active during most of support and starts activity prior to touchdown. In the terrestrial sequence, activity of extensor carpi radialis longus coincided with flexor carpi radialis. There was a period of overlap between extensor digitorum longus prior to touchdown with the other two muscles but its activity ended at or just before touchdown. Adequate recordings were not obtained from flexor digitorum superficialis in this experiment.
In experiments in which the flexors and extensors of the wrist were simultaneously recorded, there was in-phase activity of flexor carpi ulnaris, flexor carpi radialis, and extensor carpi ulnaris. Activity would commence immediately prior to touchdown and continue during the major weight-bearing portion of support phase (see Figure 2).
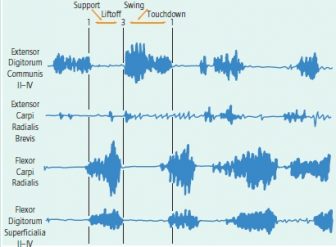
Quadrupedal walking in the hybrid male vervet. Extensor digitorum communis was active only during swing phase and was out of phase with the activity of the two flexors; its activity stopped at touchdown as in the baboon. The activity of extensor carpi radialis brevis was low, sporadic, and difficult to interpret: It may be serving to adjust the position of the hand rather than as a prime mover. Activity of flexor carpi radialis and flexor digitorum superficialis were simultaneous during support.
Activity of muscles in the palmigrade female vervet. As in the hybrid male, extensor digitorum communis and flexor digitorum superficialis were active during different phases of the step cycle. Extensor carpi radialis appeared to show two discrete bursts of low activity: one during swing phase and a smaller one at touchdown. Adequate recordings of flexor carpi radialis were not obtained in this sequence.
During trot in the male and female vervets, there were distinct bursts of activity in these muscles. At the start of activity, the hand is roughly parallel to the substrate and the fingers rectilinear to the palm. The major activity of flexor carpi ulnaris and radialis end when the proximal phalanges begin to be lifted from the substrate; at this point, the metacarpals have just passed the perpendicular in support. Activity of extensor carpi ulnaris may continue for a slightly longer period; its burst of activity appears to end when the tips of the distal phalanges are about to leave the substrate.
Activity of flexor carpi ulnaris in the baboon is initiated while the hand is outstretched during swing phase: The fingers are rectilinear to the long axis of the antebrachium. The hand and ante-brachium are moving cranially as a unit, and the hand is about to begin descent toward the substrate. Muscular activity begins at this point during both fast digitigrade walking on the ground and moderate digitigrade walking on a branch. Activity of flexor carpi radialis is similar.
In the baboon, the activity pattern of extensor carpi ulnaris often appeared to consist of two bursts separated by a short interval. These bursts occurred just at the beginning of, and near the end of, support. Major activity of the second burst stops abruptly as the metacarpals assume a perpendicular orientation relative to the substrate, rectilinear to the antebrachium. Subsequent to this, the proximal phalanges are lifted from the substrate, and flexor carpi ulnaris stops its activity. Generally, there is no activity of either extensor carpi ulnaris or flexor carpi ulnaris as the hand is lifted off the substrate and begins cranial swing.
Extensor digitorum communis is out of phase with the wrist flexors in all three animals and is active throughout most of swing. It stops its activity at touchdown, when there is a burst of activity of extensor carpi ulnaris. Full extension of the fingers in humans is produced by a combination of extensor dig-itorum and the lumbricals; the latter resist the tendency to flex at the interphalangeal joints during extension at the meta-carpophalangeal joints. Unfortunately, clear results for the long digital flexors were not obtained in the baboon. However, good recordings for both vervets clearly demonstrate that flexor digitorum superficialis and flexor digitorum profundus are in phase in their activity and are active at the same time as the wrist flexors. Both digital flexors are active during support until the basal phalanges begin to be lifted from the substrate. The muscle that is the most difficult to interpret is extensor carpi radialis. Several experiments examined this muscle in the baboon and vervets. Activity was often low and irregular. These experiments did not elucidate its function (Figures 4, 5, 6, and 7)
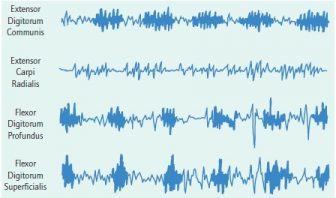
Russell Tuttle and John Basmajian conclude that flexor carpi radialis prevents extreme ulnar deviation of the wrist during knuckle-walking in apes. This muscle seems to have this function in the examined cercopithecoids: both extensor carpi ulnaris and flexor carpi ulnaris are active during support, and both would be effective ulnar deviators. It is also certainly flexing the wrist, although it is not clear how effective it is in resisting dorsiflexion because the palmigrade female vervet manifests significant activity in this muscle. Unlike results in the knuckle-walking gorilla, activity of flexor carpi ulnaris did not decrease while flexor carpi radialis remained active. Instead, flexor carpi ulnaris showed consistent activity. The result, together with my cineradiographic data on the orientation of the pisiform in digitigrade and palmigrade monkeys, suggests that flexor carpi ulnaris acts as the in-force for a first-class lever, because the muscle tendon continues as the pisohamate and pisometacarpal ligaments. Tuttle and Basmajian report little activity in extensor carpi ulnaris during knuckle-walking. In contrast, this muscle is consistently active during support in cercopithecoids. Studies by Tuttle and Basmajian and also Randall Susman and Jack Stern report relatively little activity of flexor digitorum superficialis and flexor digitorum profundus during knuckle-walking. In contrast, activity was recorded during support in both vervets. In the monkeys, extensor digitorum communis is not active during support and does not help to maintain the integrity of the joints during digitigrady in the baboon.
Simultaneous activity of flexor digitorum profundus and flexor digitorum superficialis during support in a terrestrial trot in the female vervet. Adequate recordings of extensor carpi radialis were not obtained in the experiment. Extensor digitorum coincided with the two flexors during a portion of its activity.
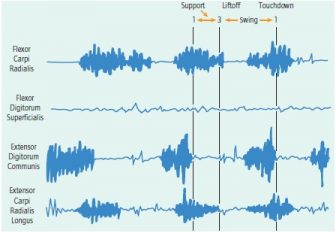
Digitigrade walk on the ground in the young male baboon. Flexor carpi radialis began its activity during the end of swing phase as the manus began its descent toward the substrate, and continued throughout most of support, ending after the antebrachium was perpendicular to the orientation of the phalanges on the substrate and the proximal phanges had begun to be lifted from the ground. Extensor digitorum communis was active only during swing phase, ending its activity at touchdown. It both extended the digits from a flexed position and maintained them in the extended state until touchdown. Adequate recordings were not obtained from the other two muscles during the experiment.
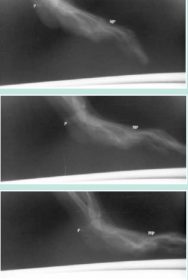
Touchdown of the hand in a palmigrade male vervet studied by cineradiography during quadruepdal walking on a treadmill. The pisiform (P) maintains a horizontal orientation throughout the sequence and is nonweight-bearing. The metacarpo-phalangeal joints (MP) are closer to the substrate prior to and at touchdown than are the proximal ends of the metacarpals. In this regard, palmigrade touchdown is more similar to digitigrade touchdown than it is to human pedal plantigrady. Source: Copyright © and courtesy of Paul F. Whitehead.
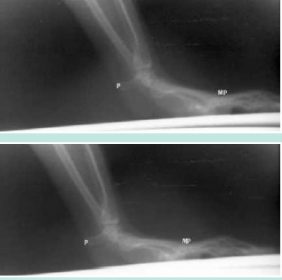
Finally, while there are still some gaps in our knowledge, there is a noticeable similarity in the muscular activity patterns of digitigrade and palmigrade cercopithecoids.
This finding is explained by cineradiographic evidence that demonstrates that the phalanges are actually inclined downward toward the substrate prior to touchdown in vervet palmigrady: This gives the hand movements greater similarity to digitigrade touchdown than had been expected. In addition, the proximal metacarpals are always higher than are the distal metacarpals during support: this is another similarity to digitigrady and a significant difference from plantigrady in bipedal humans. (See Figure 8.) The extent to which similarity in hand movement between digitigrady and palmigrady may support the hypothesis of a semiterrestrial origin of the cercopithecoids is dependent on the relative genetic conservatism of the neurological bases of hand movement.
References:
- Rose, M. D. (1994). Quadrupedalism in some Miocene catarrhines. Journal of Human Evolution 26, 387-411.
- Tuttle, R. H., & Basmajian, J. V. (1974). Electromyography of forearm musculature in Gorilla and problems related to knuckle-walking. In F. A. Jenkins Jr. (Ed.), Primate locomotion (pp. 293-347). New York: Academic Press.
- Whitehead, P. F. (1993). Aspects of the anthropoid wrist and hand. In D. L. Gebo (Ed.), Postcranial adaptation in nonhuman primates (pp. 96-120). DeKalb, IL: Northern Illinois University Press.
- Whitehead, P. F., & Larson, S. G. (1994). Shoulder motion during quadrupedal walking in Cercopithecus aethiops: Integration of cineradiographic and electromyographic data. Journal of Human Evolution 26, 525-544.
